Развитие растительного мира на Земле
По мнению ученых, планета Земля образовалась приблизительно 4,5 млрд. лет назад.
Остатки первых живых организмов датируются приблизительно 3,5 млрд. лет назад. Ранее ими считались одноклеточные гетеротрофы, пищей которым служила первичная органика. В школьной программе до сих пор указана эта точка зрения.
В настоящее время ученые склоняются к тому, что первыми живыми организмами были хемотрофные прокариоты. Они жили на дне морей и окисляли неорганические соединения, а полученную энергию использовали для синтеза органических веществ из углекислого газа.
Появление фотосинтетических пигментов, а следовательно, и способности к фотосинтезу привело к появлению автотрофных прокариот — сине-зеленых водорослей (цианей).
Следующий ароморфоз — это появление ядра. Появились автотрофные эукариоты — одноклеточные водоросли.
Далее шло появление многоклеточных организмов. Слоевище многоклеточных водорослей постепенно усложнялось, "ветвилось": происходило увеличение интенсивности фотосинтеза, а следовательно, более интенсивное образование кислорода. Постепенно формировалась азотно-кислородная атмосфера и озоновый слой.
У водорослей появилась более прогрессивная форма размножения — половое размножение, при котором начало новому поколению дает диплоидная (2n) зигота, сочетающая в себе наследственность 2-х родительских форм.
В эпоху палеозоя очертания суши постоянно изменялись, и большие участки земной поверхности то осушались, то снова затапливались океаном. Отступавшая морская вода задерживалась во впадинах. Осушение этих участков происходило постепенно. У некоторых водорослей стали появляться приспособления к обитанию вне воды. Они дали начало новой группе - псилофитам (риниофитам).

Псилофиты росли по берегам водоемов и были небольшими многоклеточными зелеными растениями. Их организм не был разделен на органы и представлял собой ветвящиеся оси, на подземных частях которых развивались ризоиды. От водорослей их отличало более сложное внутреннее строение: хорошо развита покровная ткань (кожица) и проводящие ткани (древесина и луб). Размножались псилофиты спорами, созревающими в спорангиях. Ископаемые остатки псилофитов прекрасно сохраняются в очень древних окаменелостях и являются самыми ранними известными сосудистыми растениями. Псилофиты существовали относительно недолго, уступив место папоротникам.
От псилофитов произошли высшие споровые растения: мхи, хвощи, плауны и папоротники.
Начиная с псилофитов, можно проследить 2 линии эволюции высших растений, одна из которых представлена мохообразными, а вторая — хвощами, плаунами, папоротниками, голосеменными и покрытосеменными растениями. (Существует альтернативная гипотеза, согласно которой мохообразные произошли от зеленых водорослей, параллельно с псилофитами).
Главное, что характеризует мохообразных, это преобладание в цикле их индивидуального развития гаплоидного гаметофита над диплоидным спорофитом. Гаметофит мхов — это все зеленое растение, способное к самостоятельному питанию. Спорофит мхов представлен коробочкой и полностью зависит в своем питании от гаметофита. Доминирование у мхов влаголюбивого гаметофита в условиях воздушно-наземного образа жизни оказалось нецелесообразным, поэтому мхи стали тупиковой ветвью эволюции высших растений. Этому способствовал и тот факт, что гаметофит по сравнению со спорофитом имел гаплоидный (n) набор хромосом, следовательно был более подвержен мутационным изменениям.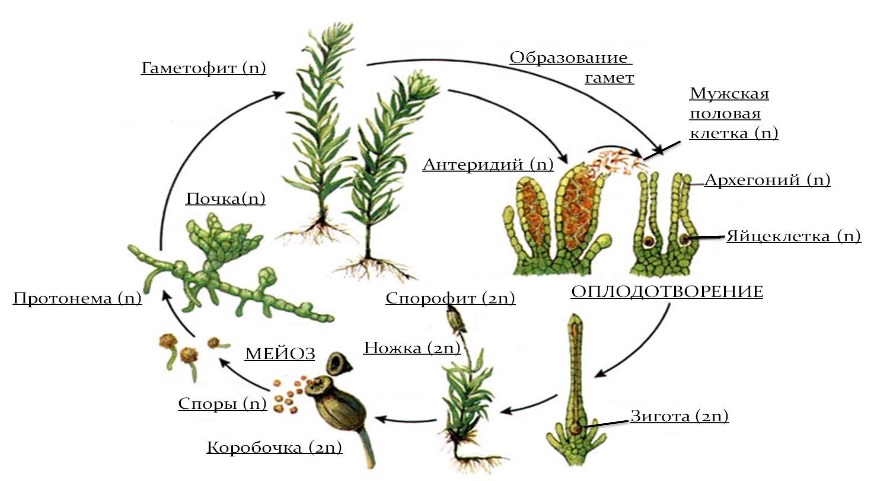
Вторая линия эволюции на пути от псилофитов к покрытосеменным представлена хвощами, плаунами, папоротниками, голосеменными и покрытосеменными растениями, в цикле индивидуального развития которых доминирует спорофит. Он представляет собой растение с корнем, стеблем, листьями, органами спороношения (у папоротников) или плодоношения (у покрытосеменных). Клетки спорофита имеют диплоидный набор хромосом, т.к. они развиваются из диплоидной зиготы. Гаметофит сильно редуцирован и приспособлен только для образования мужских и женских половых клеток. У цветковых растений женский гаметофит представлен зародышевым мешком, в котором находится яйцеклетка. Мужской гаметофит образуется при проростании пыльцы. Он состоит из одной вегетативной и одной генеративной клеток. При прорастании пыльцы из генеративной клетки возникает 2 спермия. Эти 2 мужские половые клетки участвуют в двойном оплодотворении у покрытосеменных. Оплодотворенная яйцеклетка дает начало новому поколению растения — спорофиту. Прогресс покрытосеменных обусловлен совершенствованием функции размножения.
 Древние Папортникообразные
Древние ПапортникообразныеВ это же время в палеозое расцвета достигла еще одна группа споровых растений — лепидофитов, относящихся к древним плауновидным. Это крупные (до 40 м) быстрорастущие деревья жившие не более 20 лет (точнее сказать сложно из-за отсутствия годовых колец). Лепидофиты возникли в Девонском периоде; пережили расцвет в Карбоне, когда были доминирующими растениями углеобразующих лесов; постепенно исчезали в Перми. Одни лепидофиты сильно ветвились (лепидодендроны), другие (сигиллярии) испытывали одно-два дихотомических ветвления.
 Лепидофиты
Лепидофиты
Лепидофиты имели узкие длинные листья, прираставшие к стволу и ветвям без черешка, постепенно отмирая листья опадали, но оставляли на стволе характерный рисунок, напоминающий чешую пресмыкающихся, за это они и получили свое название(греч. lepis (lepidos) — чешуя). На стволе листья располагались спирально.
Лепидофиты, хотя и выглядели как настоящие деревья, таковыми не являются, поскольку, во-первых, их ствол содержал очень мало древесины, а основной частью ствола была кора, которая не отслаивалась; во-вторых, ствол этих деревьев был зеленым, так как места прикрепления опавших листьев оставались фотосинтезирующими органами. Споры формировались в стробилах на концах ветвей. Корни, отходящие от нижней части стебля (ризофора) по спирали, были покрыты чешуевидым спиральным рисунком, соответствующим прикреплению временных придаточных корней. Корни лепидофитов находились в заболоченном грунте и не имели возможности получать кислород из грунта. Поэтому можно предположить, что газообмен у них осуществлялся, как и у полушника, через воздушные полости в корнях, куда доставлялся кислород из листьев.
Голосеменные произошли не от настоящих (типичных) папоротников, а от одной из боковых ветвей древнейших папоротниковидных растений. Эта точка зрения подтверждает правило, сформулированное известным палеонтологом Э. Коном еще в 1904 году, согласно которому в процессе эволюции новые группы организмов обычно происходят не от высших представителей предкового таксона, а от наименее специализированных его представителей.
Первыми голосеменными растениями были семенные папоротники, впоследствии полностью вымершие. Семенные папоротники были древовидными, лиановидными и травянистыми растениями. Они являлись промежуточным этапом эволюции между папоротниками и цикадовыми растениями, похожими на современные пальмы, с которыми они находятся в тесном родстве. Специальных органов для развития семян у них не было. Однако зародыш в семени был лучше защищен и снабжен питательными веществами, чем спора. Кроме того, мужские гаметы — спермии — не нуждались в присутствии воды, распространялись преимущественно ветром, а оплодотворение происходило внутри семязачатков.
 Остатки семенных папоротников
Остатки семенных папоротников
В условиях более сурового климата древние голосеменные растения постепенно вымирали, и их место занимали более приспособленные к холодным, сухим условиям древние хвойные голосеменные растения.
Первые отпечатки растений с признаками покрытосеменных обнаружены в пластах раннего мелового периода (130 млн. лет назад), а в позднем мелу покрытосеменные оказались доминирующей формой растительной жизни. Цветки покрытосеменных до такой степени разнообразны, что не существует ни одного признака или структурной особенности, которая была бы присуща всем цветкам без исключения. Тем не менее ясно, что именно появление цветка было ключевым событием в происхождении покрытосеменных. На сегодняшний день наиболее "популярной" является гипотеза о происхождении цветковых от ископаемых беннеттитовых голосеменных растений. Наиболее древней группой цветковых растений считается семейство Нимфейные (Кувшинковые) со спиральным расположением частей цветка.
Основные этапы эволюции растений
| период | таксон | новые признаки/ароморфозы | последствия |
|---|---|---|---|
3,5 млрд. лет архей | сине-зеленые водоросли (цианобактерии) | одноклеточные прокариоты; автотрофный тип питания — фотосинтез | кислородная атмосфера |
протерозой 2,6 млрд. лет | одноклеточные водоросли | появление ядра — одноклеточные эукариоты с автотрофным типом питания | разделение живых организмов на царство растений и царство животных |
протерозой (венд = эдиакарий) 600 млн. лет | многоклеточные водоросли | появление крупных форм организмов | распространение многоклеточных водорослей |
палеозой (кембрий, ордовик) 541 млн. лет | многоклеточные водоросли | усложнение строения слоевища — увеличение площади поверхности | увеличение продуктивности фотосинтеза — накопление органических отложений |
палеозой (силур) 440 млн. лет (активное изменение очертаний суши) | псилофиты | появление тканей: покровные ткани с устьицами (защита); механические ткани (опора); примитивные проводящие ткани | выход растений на сушу; переход от низших растений к высшим |
палеозой (девон) 400 млн. лет (теплый и влажный климат) | мхи папоротники, хвощи, плауны | Появление побега (лист — орган фотосинтеза). Появление проводящей ткани; развитие корневой системы; мужские гаметы со жгутиками — половой процесс зависит от воды | преимущества для жизни на суше — господство папоротников; появление лесных биоценозов; развитие почвенного покрова; увеличение количества кислорода в атмосфере |
палеозой (карбон) 360 млн. лет (похолодание и засуха) | семенные папоротники - первые голосеменные растения | появление семени: половой процесс не зависит от воды; защита и питание зародыша | предки современных голосеменных растений; |
палеозой (пермь) 370 млн. лет (засушливый климат) | голосеменные растения | развитие половых клеток во внутренних тканях; защита тканями при оплодотворении; приспособление семян к распространению ветром и водой | гибель гигантских папоротников, хвощей и плаунов в связи с засухой; быстрое завоевание суши голосеменными |
мезозой (мел) 130 млн. лет | цветковые растения | появление цветка — привлечение для опыления насекомых; усложнение строения генеративных органов: защита семяпочек завязью, а зародыша — оболочками семени и плодом; двойное оплодотворение: развитие эндосперма — питательной ткани зародыша | завоевание цветковыми растениями суши; освоение водной среды обитания |
Доказательствами эволюции растений являются палеонтологические находки их ископаемых остатков. Строматолиты (др.-греч. каменная подстилка, каменная прослойка) — ископаемые остатки цианобактериальных матов; отпечатки гигантских папоротников, хвощей, плаунов, обнаруженные в залежах каменного угля и торфяниках, многочисленные споры и пыльца в почвенных отложениях разного геологического возраста.